

Схема рефлекса растяжения физиология – 22. Мышечный тонус, его значение, природа мышечного тонуса. Рефлекторные дуги рефлекса на растяжение и сухожильного рефлекса.
Рефлексы растяжения
Мышечные веретёна и сухожильные органы Гольджиучаствуют в реализации рефлексов на растяжение, возникающих в ответ на резкое растяжение мышцы. При этом возбуждение мышечных рецепторов вызывает рефлекторное сокращение как этой, так и мышц–синергистов. На рис. 13–5 показаны рефлекторные дуги рефлексов растяжения, как моносинаптического (I), так и полисинаптического (II).
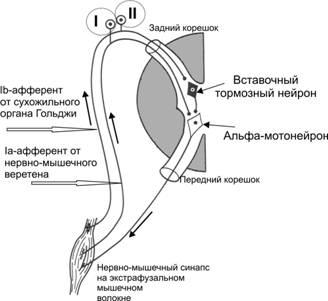
Рис.13–5.Рефлексырастяжениямоносинаптический(I, от мышечных веретён, приводит к сокращению той же мышцы)иполисинаптический(II)
Моносинаптическаядуга. I–проприоцептивные нервные волокна, отходящие от мышечного веретена, входят в задний корешок спинного мозга и сразу направляются в передний рог, где и образуют синапсы с‑мотонейронами, посылающими сигналы к мышце.
Полисинаптическаядугадополнительно включает вставочный нейрон. На рис. 13–5(II) представлена дуга тормозного рефлекса, возникающего при растяжении сухожильных рецепторовГольджи.
Динамическийистатическийрефлексырастяжения. Различают динамический и статический компоненты рефлекса растяжения.
Динамическийрефлексрастяженияосуществляется при неожиданном быстром удлинении мышцы, что приводит к столь же быстрому сокращению. Очевидно, чтофункциярефлексанаправленапротивнеожиданныхизмененийвдлинемышцы,посколькумышцасокращается,преодолеваярастяжение.
Статическийрефлексрастяжения. Динамический рефлекс растяжения совершается в доли секунды. После того, как мышца растянута до её новой длины, следует слабый статический рефлекс растяжения. Его важность состоит в том, что он продолжается в течение всего времени, пока длина мышц изменена.
Следовательно,функциястатическогорефлексарастяжениятакженаправленапротивсил,вызывающихпревышениеисходнойдлинымышцы.Сигналы, поступающие к скелетным мышцам от спинного мозга, как правило, носят дискретный характер (например, увеличить интенсивность в течение нескольких миллисекунд, изменить уровень интенсивности, уменьшить силу сокращения и т.д.). То обстоятельство, что в норме даже самые быстрые движения осуществляются плавно, как раз и связано с наличием динамического и статического компонентов рефлексов растяжения. Другими словами, динамическийистатическийкомпонентырефлексарастяжения
Участие мышечных веретён в произвольных движениях
Сигналы из моторной коры и других областей мозга, поступающие к ‑мотонейронам спинного мозга, одновременно возбуждают и‑мотонейроны (феномен коактивации– и‑мотонейронов). 31% эфферентов к скелетным мышцам составляют нервные волокна типа A. В результатеприкаждоммышечномсокращениипроисходитодновременноесокращениеиэкстра–иинтрафузальныхМВ.
-Эфферентная система активируется импульсами, поступающими из бульборетикулярной активирующей формации мозгового ствола, и опосредованно — импульсами, поступающими в бульборетикулярную активирующую формацию из мозжечка, базальных ганглиев, коры больших полушарий. Это связано с тем, что бульборетикулярная активирующая формация непосредственно связана с антигравитационными сокращениями, и антигравитационные мышцы имеют самую высокую концентрацию мышечных веретён.
Поэтому–эфферентныйдемпфирующиймеханизмособеннопроявляетсявовремяходьбыибега. Другаяважнаяфункциясистемымышечныхверетёнзаключаетсявстабилизацииположениятелавовремянапряжённоймышечнойдеятельности. Физиологический механизм этого эффекта состоит в том, что при динамической работе (одновременно с сокращением групп сгибательных мышц) усиливаются рефлексы растяжения мышц разгибательной группы. Любое усиление сокращения с одной стороны сустава демпфируется усилением рефлексов растяжения с другой стороны. В результате достигается стабилизация положения сустава.
studfile.net
Физиология растягивания
Мышечные волокна
Скелетные мышцы состоят из пучков мышечных волокон. Мышечные волокна могут сокращаться, расслабляться или удлиняться.
Мышечные волокна сокращаются под действием электрического импульса, поступающего от нерва. Одно мышечное волокно всегда сокращается полностью, создавая фиксированную силу. Сила, развиваемая всей мышцей, определяется количеством волокон, вовлеченных в это действие нервной системой.
Соединительные ткани
Соединительные ткани в опорно-двигательном аппарате — это связки, сухожилия и фасции. Связки обоими концами крепятся к костям сустава, сухожилия крепят мышцы к костям, а фасции обволакивают группы мышц и пучки мышечных волокон.
Связки и сухожилия содержат большое количество белка коллагена, который придает им жесткость. Связки и сухожилия практически не растягиваются — избыточная нагрузка на эти ткани может привести к травме. Фасции относятся к эластичным соединительным тканям (содержат белок эластин), и их растягивание повышает гибкость. Большинство упражнений на гибкость направлено на растяжение фасций.
Механизм растягивания
При растягивании сначала вытягиваются мышечные волокна, а затем в направлении вытягивающего усилия выравниваются коллагеновые волокна соединительной ткани. Это позволяет упорядочить волокна в направлении растяжения, благодаря чему в упражнениях на растяжение восстанавливается здоровая структура ткани.
При растяжении мышц часть мышечных волокон растягивается, а часть остается в исходном состоянии. Длина мышцы определяется количеством растянутых волокон (аналогично тому, как длина и сила сжатой мышцы определяется количеством сжатых волокон). Чем больше растянутых волокон, тем длиннее растянутая мышца.
Рефлекс растяжения
В толще мышц находятся нервно-мышечные веретена — нервные окончания, реагирующие на удлинение мышц. Веретена располагаются параллельно мышечным волокнам и растягиваются вместе с ними. При этом они регистрируют как изменение длины мышцы, так и скорость этого изменения.
При сильном или резком удлинении мышцы сигнал от веретен вызывает защитный рефлекс растяжения — мышца самопроизвольно сокращается в попытке помешать удлинению.
Рефлекс растяжения имеет статическую и динамическую составляющие. Статическая составляющая сохраняется все время, пока мышца растянута. Динамическая составляющая — это ответ на скорость изменения длины мышцы, он может быть очень сильным при резком удлинении мышцы и снижается, когда скорость удлинения мышцы уменьшается.
Одна из целей удержания позы растяжения в течение некоторого времени — дать мышечным веретенам адаптироваться к новой длине мышцы с уменьшением интенсивности их сигналов в новом положении.
Реакция удлинения
При сокращении мышца создает натяжение на сухожилии, к которому она крепится. На это реагирует сухожильный орган Гольджи — тип нервного окончания, который находится в местах соединения волокон мышц и сухожилий. Сухожильный орган реагирует на величину возникающей силы натяжения (статический эффект) и на скорость изменения этой силы (динамический эффект: резкое натяжение вызывает сильный сигнал).
Когда сигнал от сухожильного органа превосходит определенный порог, возникает защитная реакция удлинения — резкое самопроизвольное расслабление всей мышцы.
Вторая причина для удержания позы растяжения в течение некоторого времени — позволить произойти реакции удлинения, которая помогает мышце расслабиться (преодолевая рефлекс сжатия).
Взаимное торможение
По отношению к выполняемому движению мышцы можно отнести к одному из четырех типов:
- Агонисты — вызывают движение.
- Антагонисты — вызывают противоположное движение; отвечают за возвращение в начальное положение тела.
- Синергисты — корректируют действие мышцы-агониста для обеспечения нужного направления результирующей силы.
- Стабилизаторы — удерживают остальную часть тела при выполнении движения.
Агонисты и антагонисты обычно находятся с противоположных сторон сустава (например, бицепс и трицепс), а синергисты — на той же стороне, что и агонисты, рядом с ними. При работе крупных мышц-агонистов часто вовлекаются в качестве синергистов находящиеся рядом меньшие мышцы.
Сокращение мышцы-агониста может привести к рефлекторному расслаблению ее антагониста. Это явление называется взаимным торможением.
Взаимное торможение происходит не при всех движениях. Иногда возникает явление совместного сжатия. Например, при приседаниях сжимаются как мышцы брюшного пресса, так и их антагонисты — разгибатели спины.
Упражнения на растяжения легче выполнять с расслабленными, а не с сокращающимися мышцами. Для этого можно использовать эффект взаимного торможения (когда он происходит) — заставляя мышцы-антагонисты расслабляться за счет сжатия агонистов. Также желательно расслабить синергистов мышцы, которую вы растягиваете. Например, при растягивании икроножной мышцы, следует нагнуть стопу, сжав мышцы передней поверхности голени. С другой стороны, икроножная мышца — синергист подколенного сухожилия, поэтому желательно расслабить и его. Для этого нужно выпрямить ногу, сжав тем самым антагониста подколенного сухожилия — четырёхглавую мышцу бедра.
mhlife.ru
Клиническое значение рефлексов растяжения
В повседневной клинической практике рефлексы растяжения используют для определения состояния уровня возбуждения, или «тонуса» ЦНС. На рис. 13–6 приведены некоторые рефлексы спинного мозга. Быстрое растяжение мышцы механическим ударом по её сухожилию вызывает сокращение всей мышцы. Например, легкий удар по связке надколенника (рис. 13–6А)вызывает динамический рефлекс растяжения — сокращение мышц бедра и разгибание голени.
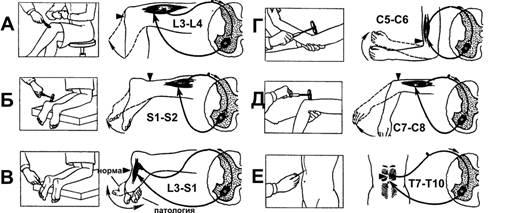
Рис.13–6.Рефлексыспинногомозга.А—коленныйрефлекс;Б—ахилловрефлекс;В—подошвенныйрефлекс (рефлекс Бабинского);Г—сгибательныйрефлекс предплечья;Д—разгибательныйрефлекс предплечья;Е—брюшнойрефлекс.
Дуга моносинаптического коленного рефлекса такова: мышечные веретёна четырехглавой мышцы бедра спинномозговой узелзадние корешкизадние рога III поясничного сегмента‑мотонейроны передних рогов того же сегментаэкстрафузальные МВ четырехглавой мышцы бедра. Подобные рефлексы могут быть получены почти у всех мышц тела. Усиление рефлексов растяжения расценивается как повышение облегчающих, активирующих влияний верхних уровней ЦНС. Напротив, депрессия облегчающих влияний со стороны ЦНС ослабляет рефлексы растяжения. Эти рефлексы применяют для оценки наличия или отсутствия спастичности мышц. Так, при инсульте или опухоли мозга наблюдается значительное усиление рефлексов растяжения на поражённой стороне тела.
Функции рецепторов Гольджи
Сухожильный орган Гольджи(см. рис. 7–9 и 13–3Б [в книге]) регистрирует напряжение, развиваемое МВ в области прикрепления к сухожилию. Таким образом,мышечныеверетенаопределяютдлинумышцыиизмененияеёдлины, асухожильныеорганы—мышечноенапряжение. Подобно мышечным веретёнам, сухожильный орган отвечает на быстрое увеличение мышечного напряжения динамической реакцией, а на медленные изменения напряжения — статической реакцией. Сигналы от сухожильного органа идут по миелиновым и быстропроводящим волокнам I. Сигналы поступают в задний рог соответствующего сегмента спинного мозга (рис. 13–5,II) и полисинаптически (через тормозный вставочный нейрон) переключаются на‑мотонейроны. Кроме того, информация от сухожильных органов передаётся по спинномозжечковому тракту, а также по другим афферентным путям, идущим к коре больших полушарий.
Аутогенноеторможение. Так как сухожильный органГольджи(в отличие от мышечных веретён) связан с экстрафузальными МВ, он реагирует как на растяжение, так и на сокращение мышцы. Сильное быстрое растяжение вызывает интенсивное сокращение. В тот момент, когда напряжение, развиваемое мышцей, становится достаточно высоким, сокращение мышцы внезапно прекращается, и она расслабляется. Это расслабление в ответ на сильное растяжение —возвратныйрефлексрастяжения,илиаутогенноеторможение. Такого же рода тормозное влияние рецептор Гольджи оказывает на мышцу в момент сильного сокращения.
Такимобразом,органГольджифункционируеткакпреобразовательвцепиобратнойсвязи,регулируясилумышечногосокращения.
Полисинаптические рефлексы
Сгибательныйрефлексвозникает в ответ на сильные раздражения болевых рецепторов конечности и заключается в сокращении сгибательных мышц с одновременным расслаблением разгибательных мышц, чтобы удалить конечность от источника раздражения (рис. 13–7). Болевое раздражение, наносимое на какой-либо участок тела, также вызывает реакцию ухода от раздражения и носит более общее название —рефлексотдёргивания.
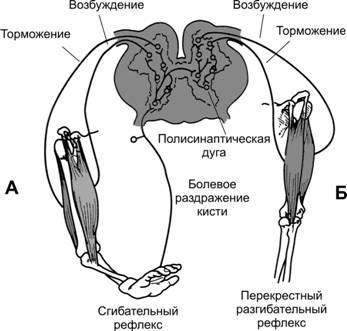
Рис.13–7.Реципрокноеторможение.А— сгибательный рефлекс.Б— перекрестный разгибательный рефлекс.
На рис. 13–7А показаны пути осуществления сгибательного рефлекса в ответ на болевой стимул, приложенный к кисти. Импульсы от болевого раздражения кисти поступают в пул интернейронов, где взаимодействуют с большим числом вставочных нейронов, активируя несколько нейронных цепей до того, как сигнал достигнет мотонейронов. Необходима активация минимально трёх нейронных цепей: дивергенция импульсов для вовлечения различных мышц в рефлекс отдергиванияторможение антагонистических мышц —реципрокноеторможениеследовые разряды после действия стимула.
Перекрестныйразгибательныйрефлекс. Возникновение болевого сгибательного рефлекса одной конечности через 200–500 мс вызывает разгибание конечности на противоположной стороне. Это перекрестный разгибательный рефлекс. Разгибание противоположной конечности направлено на оттягивание тела от источника боли, вызвавшего рефлекс отдергивания. На рис. 13–7Б показаны пути реализации перекрестного разгибательного рефлекса. Из нейронного пула сгибательного рефлекса сигналы поступают на противоположную сторону спинного мозга и интегрируются здесь со вставочными нейронами для осуществления реципрокного разгибательного рефлекса.
Реципрокныевзаимоотношения— те случаи, когда возбуждение одной группы мышц тормозит другую. Например, рефлекс растяжения одной мышцы часто одновременно тормозит антагонистические мышцы. Это явление —феноменреципрокноготорможения, а иннервация, осуществляющая эти взаимоотношения, называетсяреципрокнойиннервацией. Подобные же отношения существуют между мускулатурой двух сторон тела.
Постуральныерефлексыирефлексыдвижения. Постуральные (позотонические, статические) рефлексы обеспечивают поддержание в пространстве определённого положения всего тела или его части (например, конечности). Так, давление на подушечки стопы спинального животного вызывает реакцию вытягивания конечности, направленного против давления. Этоположительнаяподдерживающаяреакция. Если спинальных животных (с перерезкой шейного отдела спинного мозга ниже уровня С4) укладывать на бок, они производят ряд некоординированных движений для восстановления положения стоя. Указанные явления получили названиеспинномозговойвращательныйрефлекс. Спинальные животные совершаютритмическиешагательныедвижения. Это свидетельствует о существовании на уровне спинного мозга закреплённых нейронных пулов, осуществляющих рефлекторный сложнокоординированный акт ходьбы. В табл. 13–2 приведены основные постуральные рефлексы.
Таблица 13–2. Важнейшие постуральные рефлексы
Рефлекс | Стимулы | Ответ | Рецепторы | Уровень |
Растяжения | Растяжение мышцы | Сокращение мышцы | Мышечные веретена | Спинной мозг Продолговатый мозг |
Положительная поддерживающая реакция | Контакт с ладонью или подошвой | Конечность вытягивается для поддержания тела | Проприорецепторы дистальных сгибателей | Спинной мозг Продолговатый мозг |
Отрицательная поддерживающая реакция | Растяжение мышцы | Снятие положительной поддерживающей реакции | Проприорецепторы в разгибателях | Спинной мозг |
Тонические лабиринтные | Гравитация | Сокращение разгибателей конечностей | Отолитовые органы | Продолговатый мозг |
Тонические шейные | Поворот головы | Изменение паттерна сокращения разгибателей | Проприорецепторы шеи | Продолговатый мозг |
Лабиринтные выпрямительные | Гравитация | Удержание положения головы | Отолитовые органы | Средний мозг |
С тела на выпрямление головы | Давление на сторону тела | Выпрямление головы | Экстерорецепторы кожи | Средний мозг |
Шейные выпрямительные | Растяжение шейных мышц | Выпрямление туловища и плеч | Мышечные веретена | Средний мозг |
С тела на выпрямление тела | Давление на сторону тела | Выпрямление тела | Экстерорецепторы кожи | Средний мозг |
Зрительные выпрямительные | Зрительные | Выпрямление головы | Фоторецепторы глаза | Кора мозга |
Установочные реакции | Зрительные, экстероцептивные, проприоцептивные | Конечности располагаются в позиции поддержания тела | Фоторецепторы Экстерорецепторы Проприорецепторы | Кора мозга |
Реакции подпрыгивания | Смещение тела вбок при стоянии | Отпрыгивание, поддержание конечностей в позиции поддержки тела | Мышечные веретена | Кора мозга |
Постуральные рефлексы, приведённые в табл. 13–2, показывают, что на уровне спинного, продолговатого и среднего мозга осуществляются сложнокоординированные реакции позы и движений. Автоматизм этих реакций разгружает более высокие уровни регуляции мышечной активности от управления стандартными положениями частей тела по отношению друг к другу.
studfile.net
Клиническое значение рефлексов растяжения
В повседневной клинической практике рефлексы растяжения используют для определения состояния уровня возбуждения, или «тонуса» ЦНС. На рис. 13–6 приведены некоторые рефлексы спинного мозга. Быстрое растяжение мышцы механическим ударом по её сухожилию вызывает сокращение всей мышцы. Например, легкий удар по связке надколенника (рис. 13–6А)вызывает динамический рефлекс растяжения — сокращение мышц бедра и разгибание голени.
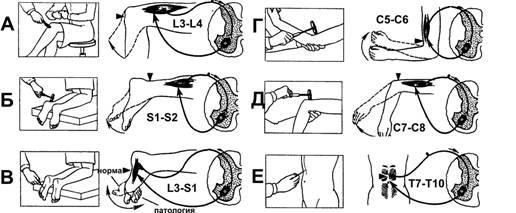
Рис.13–6.Рефлексыспинногомозга.А—коленныйрефлекс;Б—ахилловрефлекс;В—подошвенныйрефлекс (рефлекс Бабинского);Г—сгибательныйрефлекс предплечья;Д—разгибательныйрефлекс предплечья;Е—брюшнойрефлекс.
Дуга моносинаптического коленного рефлекса такова: мышечные веретёна четырехглавой мышцы бедра спинномозговой узелзадние корешкизадние рога III поясничного сегмента‑мотонейроны передних рогов того же сегментаэкстрафузальные МВ четырехглавой мышцы бедра. Подобные рефлексы могут быть получены почти у всех мышц тела. Усиление рефлексов растяжения расценивается как повышение облегчающих, активирующих влияний верхних уровней ЦНС. Напротив, депрессия облегчающих влияний со стороны ЦНС ослабляет рефлексы растяжения. Эти рефлексы применяют для оценки наличия или отсутствия спастичности мышц. Так, при инсульте или опухоли мозга наблюдается значительное усиление рефлексов растяжения на поражённой стороне тела.
Функции рецепторов Гольджи
Сухожильный орган Гольджи(см. рис. 7–9 и 13–3Б [в книге]) регистрирует напряжение, развиваемое МВ в области прикрепления к сухожилию. Таким образом,мышечныеверетенаопределяютдлинумышцыиизмененияеёдлины, асухожильныеорганы—мышечноенапряжение. Подобно мышечным веретёнам, сухожильный орган отвечает на быстрое увеличение мышечного напряжения динамической реакцией, а на медленные изменения напряжения — статической реакцией. Сигналы от сухожильного органа идут по миелиновым и быстропроводящим волокнам I. Сигналы поступают в задний рог соответствующего сегмента спинного мозга (рис. 13–5,II) и полисинаптически (через тормозный вставочный нейрон) переключаются на‑мотонейроны. Кроме того, информация от сухожильных органов передаётся по спинномозжечковому тракту, а также по другим афферентным путям, идущим к коре больших полушарий.
Аутогенноеторможение. Так как сухожильный органГольджи(в отличие от мышечных веретён) связан с экстрафузальными МВ, он реагирует как на растяжение, так и на сокращение мышцы. Сильное быстрое растяжение вызывает интенсивное сокращение. В тот момент, когда напряжение, развиваемое мышцей, становится достаточно высоким, сокращение мышцы внезапно прекращается, и она расслабляется. Это расслабление в ответ на сильное растяжение —возвратныйрефлексрастяжения,илиаутогенноеторможение. Такого же рода тормозное влияние рецептор Гольджи оказывает на мышцу в момент сильного сокращения.
Такимобразом,органГольджифункционируеткакпреобразовательвцепиобратнойсвязи,регулируясилумышечногосокращения.
Полисинаптические рефлексы
Сгибательныйрефлексвозникает в ответ на сильные раздражения болевых рецепторов конечности и заключается в сокращении сгибательных мышц с одновременным расслаблением разгибательных мышц, чтобы удалить конечность от источника раздражения (рис. 13–7). Болевое раздражение, наносимое на какой-либо участок тела, также вызывает реакцию ухода от раздражения и носит более общее название —рефлексотдёргивания.
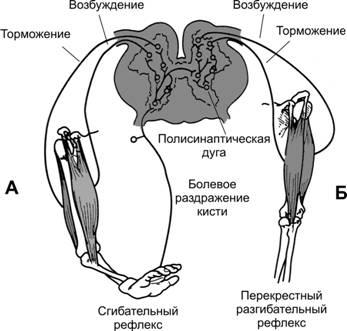
Рис.13–7.Реципрокноеторможение.А— сгибательный рефлекс.Б— перекрестный разгибательный рефлекс.
На рис. 13–7А показаны пути осуществления сгибательного рефлекса в ответ на болевой стимул, приложенный к кисти. Импульсы от болевого раздражения кисти поступают в пул интернейронов, где взаимодействуют с большим числом вставочных нейронов, активируя несколько нейронных цепей до того, как сигнал достигнет мотонейронов. Необходима активация минимально трёх нейронных цепей: дивергенция импульсов для вовлечения различных мышц в рефлекс отдергиванияторможение антагонистических мышц —реципрокноеторможениеследовые разряды после действия стимула.
Перекрестныйразгибательныйрефлекс. Возникновение болевого сгибательного рефлекса одной конечности через 200–500 мс вызывает разгибание конечности на противоположной стороне. Это перекрестный разгибательный рефлекс. Разгибание противоположной конечности направлено на оттягивание тела от источника боли, вызвавшего рефлекс отдергивания. На рис. 13–7Б показаны пути реализации перекрестного разгибательного рефлекса. Из нейронного пула сгибательного рефлекса сигналы поступают на противоположную сторону спинного мозга и интегрируются здесь со вставочными нейронами для осуществления реципрокного разгибательного рефлекса.
Реципрокныевзаимоотношения— те случаи, когда возбуждение одной группы мышц тормозит другую. Например, рефлекс растяжения одной мышцы часто одновременно тормозит антагонистические мышцы. Это явление —феноменреципрокноготорможения, а иннервация, осуществляющая эти взаимоотношения, называетсяреципрокнойиннервацией. Подобные же отношения существуют между мускулатурой двух сторон тела.
Постуральныерефлексыирефлексыдвижения. Постуральные (позотонические, статические) рефлексы обеспечивают поддержание в пространстве определённого положения всего тела или его части (например, конечности). Так, давление на подушечки стопы спинального животного вызывает реакцию вытягивания конечности, направленного против давления. Этоположительнаяподдерживающаяреакция. Если спинальных животных (с перерезкой шейного отдела спинного мозга ниже уровня С4) укладывать на бок, они производят ряд некоординированных движений для восстановления положения стоя. Указанные явления получили названиеспинномозговойвращательныйрефлекс. Спинальные животные совершаютритмическиешагательныедвижения. Это свидетельствует о существовании на уровне спинного мозга закреплённых нейронных пулов, осуществляющих рефлекторный сложнокоординированный акт ходьбы. В табл. 13–2 приведены основные постуральные рефлексы.
Таблица 13–2. Важнейшие постуральные рефлексы
Рефлекс | Стимулы | Ответ | Рецепторы | Уровень |
Растяжения | Растяжение мышцы | Сокращение мышцы | Мышечные веретена | Спинной мозг Продолговатый мозг |
Положительная поддерживающая реакция | Контакт с ладонью или подошвой | Конечность вытягивается для поддержания тела | Проприорецепторы дистальных сгибателей | Спинной мозг Продолговатый мозг |
Отрицательная поддерживающая реакция | Растяжение мышцы | Снятие положительной поддерживающей реакции | Проприорецепторы в разгибателях | Спинной мозг |
Тонические лабиринтные | Гравитация | Сокращение разгибателей конечностей | Отолитовые органы | Продолговатый мозг |
Тонические шейные | Поворот головы | Изменение паттерна сокращения разгибателей | Проприорецепторы шеи | Продолговатый мозг |
Лабиринтные выпрямительные | Гравитация | Удержание положения головы | Отолитовые органы | Средний мозг |
С тела на выпрямление головы | Давление на сторону тела | Выпрямление головы | Экстерорецепторы кожи | Средний мозг |
Шейные выпрямительные | Растяжение шейных мышц | Выпрямление туловища и плеч | Мышечные веретена | Средний мозг |
С тела на выпрямление тела | Давление на сторону тела | Выпрямление тела | Экстерорецепторы кожи | Средний мозг |
Зрительные выпрямительные | Зрительные | Выпрямление головы | Фоторецепторы глаза | Кора мозга |
Установочные реакции | Зрительные, экстероцептивные, проприоцептивные | Конечности располагаются в позиции поддержания тела | Фоторецепторы Экстерорецепторы Проприорецепторы | Кора мозга |
Реакции подпрыгивания | Смещение тела вбок при стоянии | Отпрыгивание, поддержание конечностей в позиции поддержки тела | Мышечные веретена | Кора мозга |
Постуральные рефлексы, приведённые в табл. 13–2, показывают, что на уровне спинного, продолговатого и среднего мозга осуществляются сложнокоординированные реакции позы и движений. Автоматизм этих реакций разгружает более высокие уровни регуляции мышечной активности от управления стандартными положениями частей тела по отношению друг к другу.
studfile.net
Рефлексы, их классификация.
Рефлекс (от лат. reflexus — отражённый) — стереотипная реакция живого организма на раздражитель, проходящая с участием нервной системы. Рефлексы существуют у многоклеточных живых организмов, обладающих нервной системой, осуществляются посредством рефлекторной дуги.] Рефлекс — основная форма деятельности нервной системы.Термин «рефлекс» был введен французским ученым Р. Декартом в XVII в. Но для объяснения психической деятельности он был применен основоположником русской материалистической физиологии И. М. Сеченовым.
Предположение о полностью рефлекторном характере деятельности высших отделов головного мозга впервые было развито ученым-физиологом И. М. Сеченовым. До него физиологи и неврологи не решались поставить вопрос о возможности физиологического анализа психических процессов, которые предоставлялось решать психологии.
Далее идеи И. М. Сеченова получили развитие в трудах И. П. Павлова, который открыл пути объективного экспериментального исследования функций коры, разработал метод выработки условных рефлексов и создал учение о высшей нервной деятельности. Павлов в своих трудах ввел деление рефлексов на безусловные, которые осуществляются врожденными, наследственно закрепленными нервными путями, и условные, которые, согласно взглядам Павлова, осуществляются посредством нервных связей, формирующихся в процессе индивидуальной жизни человека или животного.
Большой вклад в формирование учения о рефлексах внёс Чарлз С. Шеррингтон (Нобелевская премия по физиологии и медицине, 1932). Он открыл координацию, взаимное ингибирование и облегчение рефлексов.
Классификация
По ряду признаков рефлексы могут быть разделены на группы[3].
По типу образования: условные и безусловные рефлексы.
По видам рецепторов: экстероцептивные (кожные, зрительные, слуховые, обонятельные), интероцептивные (с рецепторов внутренних органов) и проприоцептивные (с рецепторов мышц, сухожилий, суставов)
По эффекторам: соматические, или двигательные (рефлексы скелетных мышц), например флексорные, экстензорные, локомоторные, статокинетические и др.; вегетативные внутренних органов — пищеварительные, сердечно-сосудистые, выделительные, секреторные и др.
По биологической значимости: оборонительные, или защитные, пищеварительные, половые, ориентировочные.
По степени сложности нейронной организации рефлекторных дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также один или несколько промежуточных нейронов и имеют два или несколько синаптических переключений (например, флексорный).
По характеру влияний на деятельность эффектора: возбудительные — вызывающие и усиливающие (облегчающие) его деятельность, тормозные — ослабляющие и подавляющие её (например, рефлекторное учащение сердечного ритма симпатическим нервом и урежение его или остановка сердца — блуждающим).
По анатомическому расположению центральной части рефлекторных дуг различают спинальные рефлексы и рефлексы головного мозга. В осуществлении спинальных рефлексов участвуют нейроны, расположенные в спинном мозге. Пример простейшего спинального рефлекса — отдергивание руки от острой булавки. Рефлексы головного мозга осуществляются при участии нейронов головного мозга. Среди них различают бульбарные, осуществляемые при участии нейронов продолговатого мозга; мезэнцефальные — с участием нейронов среднего мозга; кортикальные — с участием нейронов коры больших полушарий головного мозга.
studfile.net
ФИЗИОЛОГИЯ ДВИЖЕНИЙ
передние конечности разгибаются. Дальнейшее прямолинейное смещение головы в пространстве при свободном падении вызывает разгибание задних конечностей. Последнее можно объяснить лабиринтным рефлексом на падение (линейное ускорение). Благодаря совокупному действию этих рефлексов голова и тело кошки принимают нормальное положение, конечности тонически разгибаются и готовы принять вес тела, когда животное достигнет земли.
Таким образом, положение тела и головы в пространстве и по отношению друг к другу принадлежит к наиболее застрахованным и автоматически выполняемым реакциям ЦНС. Благодаря взаимной деятельности большого количества разнородных рефлексов осуществляется единая конечная цель – правильная установка головы и тела в пространстве. Статокинетические и статические реакции взаимно дополняют друг друга: благодаря кинетическим реакциям выполняется движение, приводящее отдельные части тела в такое положение, в котором они потом удерживаются статическими рефлексами.
Глазодвигательная система млекопитающих. В эволюции наземных позвоночных животных возникла необходимость сохранять неподвижность изображения на сетчатке не только в состоянии покоя или медленных движений, но также во время локомоции. Эволюция глазодвигательной системы у этих животных происходила в тесном взаимодействии с эволюцией зрительной системы. С точки зрения эволюции глазодвигательной системы всех позвоночных животных можно условно разделить на две группы: «фовеальные» животные, сетчатка которых имеет fovea – и они обладают достаточно острым зрением, и «афовеальные» животные – с недостаточно острым зрением. У «фовеальных» животных (к ним относится и человек) глаза расположены во фронтальной плоскости, у них есть бинокулярное зрение. Зрительная ямка, или fovea,-это небольшая область в сетчатке с повышенной плотностью фоторецепторов, причем только в этой области находятся колбочки – фоторецепторы дневного (и цветового) зрения. Глазодвигательная система млекопитающих, которая сформировалась как необходимое дополнение зрительной системы, позволяет рассматривать зрительный мир. С точки зрения физиологии непрерывно решается задача – движением глаз поместить на зрительную ямку наиболее значимые части изображения. Что будет, если отключить fovea? Это происходит с каждым из нас при снижении уровня освещенности, например, вечером. Известно, что мы в этих условиях не видим мелких предметов, не можем читать. Для такого зрения движения глаз не обязательны. Действительно, у животных с невыраженной fovea, например собаки, коровы, быстрые движения глаз отсутствуют.
Глазодвигательная система млекопитающих выполняет следующие задачи: 1) сохраняет неподвижным изображение внешнего мира на сетчатке во время движения животного относительно этого мира; 2) выделяет во внешнем мире некоторые объекты, помещает их в зоне сетчатки с высоким разрешением (например, fovea у человека) и прослеживает их движениями глаз и головы; 3) осуществляет скачкообразные (саккадические) перемещения взора для сканирования (рассматривания) внешнего мира. «Афовеальные» животные, например кошка, собака, кролик и др., не заботятся о положении изображения на сетчатке, и вестибулярная и оптокинетическая системы только предотвращают скольжение изображения по сетчатке. У человека в связи с изложенными выше требованиями, кроме вестибулоокулярной и оптокинетической систем, имеется также система прослеживания, вергенции и саккад. У него развились также рефлекторные механизмы содружественного движения глаз и головы.
Саккадная система в полной мере развита только у обезьян и человека. Именно у них есть бинокулярное зрение, и есть «фовеальное» зрение.
Саккады – это быстрые содружественные отклонения глаз в быструю фазу вестибулярного и оптомоторного нистагмов, начальная фаза реакции прослеживания, когда скачком глаза «захватывается» движущаяся зрительная цель при зрительном обследовании внешнего мира. Структуры мозга, обеспечивающие быстрые, скачкообразные движения глаз – саккады, находятся в ретикулярной формации мозгового ствола. Однако высший центр произвольного управления движениями глаз локализован в коре лобной доли мозга и называется
фронтальным глазодвигательным полем (8-е и 10-е поля).
Активность нейронов мозгового ствола. Схема, представленная на рис. 5.15, показывает связи и вероятные события, приводящие к саккаде глаза. Важными элементами этой схемы являются пачечные нейроны и нейроны-паузы. Во время фиксации взора клетки-паузы
studfile.net
ОБРАТНЫЙ РЕФЛЕКС РАСТЯЖЕНИЯ МЫШЦ
В мышцах также есть сухожильные веретена, которые задействованы во время активного и пассивного мышечного сокращения. Когда напряжение в мышце слишком большое, то есть имеется вероятность ее разрыва, срабатывает обратный рефлекс растяжения мышцы, чтобы эта мышца расслабилась.
В качестве примера мы можем сказать, что обратный рефлекс растяжения мышцы срабатывает, когда происходит сильное мышечное сокращение. Цель такого рефлекса – защитить мышцу и области ее прикрепления от возможной травмы (растяжение, разрыв, разрыв волокон), случившейся в результате большой перегрузки.
Именно механизм обратного растяжения мышцы делает растяжку НВП более эффективной.
Оба эти рефлекса – рефлекс растяжения мышцы и обратный рефлекс растяжения мышцы – определенным образом контролируют движения.
ЧАРЛЬЗ СКОТТ ШЕРРИНГТОН (1857-1952)
Закон Шеррингтона: Когда мышца получает нервный импульс к сокращению, ее антагонист одновременно получает другой импульс к расслаблению (реципрокная иннервация).
ВИДЫ РАСТЯЖКИ
Так же как существуют различные виды растяжки, есть различные виды упражнений, помогающие увеличить амплитуду движения в суставе.
ПАССИВНАЯ РАСТЯЖКА
Пассивная растяжка заключается в принятии какой-либо позы и ее удержании при помощи другой части тела, другого человека или какого-либо оборудования. Тот, кого растягивают, должен быть расслаблен, а тот, кто вытягивает, направляет вытягиваемую конечность, чтобы увеличить амплитуду растяжки. Например, подъем ноги из положения стоя и удержание ее в этом положении при помощи рук. Такая техника растяжки называется пассивной, потому что вытягивание происходит пассивным образом. Такой способ растяжки популярен среди гимнастов, у которых максимальная гибкость необходима как раз в конечной точке амплитуды.
К сожалению, если такая растяжка выполняется кем-то, кто не знает о возможной опасности и не полностью концентрируется на цели данной растяжки, это может привести к травме. Очень важно получать обратную связь от человека, которого вытягивают, чтобы ответственность лежала именно на нем. Пассивная растяжка применяется в реабилитации, когда активная растяжка причиняет боль.
АКТИВНАЯ РАСТЯЖКА
Активная растяжка – это когда человек выполняет движения на растяжку самостоятельно. Этот вид растяжки считается более безопасным, так как человек, выполняющий растяжку, понимает, что он делает и с меньшей вероятностью приничинит себе травму. Такая растяжка заключается в принятии и удержании какой-либо позы без чьей-либо помощи, разве что мышцы-агониста. Например, поъем ноги из положения стоя и ее удержание без какой-либо помощи. Амплитуда, которая достигается при такой растяжке, является результатом мышечной силы, а не вытяжения мышц-антагонистов. Напряжение мышцы-агониста скорее поможет мышцам-антагонистам расслабиться в результате реципрокного торможения. Некоторые позы в йоге являются активными растяжками.
БАЛЛИСТИЧЕСКАЯ РАСТЯЖКА
Также называется «пружинящая растяжка». В этом виде растяжки используется инерция части тела, чтобы вывести сустав на пределы свой нормальной амплитуды движения. В основном, такая растяжка выполняется активным способом, но может выполняться и пассивно, то есть с помощью. Обычно в спорте предпочитают именно этот вид растяжки, так как считается, что пружинящие движения активизируют рефлекс растяжения мышцы, поскольку этот рефлекс защищает мышцу и сустав от травмы, которая может привести к укорачиванию мыщцы, а не к ее удлинению. Пружинящие движения не позволяют произойти обратному рефлексу растяжения мышцы и позволяют мышце расслабиться и вытянуться.
Обычно не используется как вид растяжки из соображений безопасности, так как есть вероятность перерастяжения мышцы во время пружинящего движения. Исключением будут виды спорта, в которых требуется баллистическая растяжка. Например, там, где есть кики ногами с полной амплитудой движения, как в некоторых видах боевых искусств или ритмической гимнастике.
Этот вид растяжки распространен среди танцоров и выполняется с ритмическими пружинящими движениями, чтобы заставить нужную мышцу вытянуться.
ДИНАМИЧЕСКАЯ РАСТЯЖКА
Динамическую растяжку не следует путать с баллистической. Несмотря на то, что действия в обоих видах растяжки похожи, скорость выполнения динамической растяжки находится под контролем, чтобы сустав мог плавно достичь конечтой точки своей амплитуды движения. Динамическая растяжка начинается неагрессивно. Такая растяжка помогает улучшить динамическую гибкость и очень хороша для разминки перед основной тренировкой.
СТАТИЧЕСКАЯ РАСТЯЖКА
Статическая растяжка – это когда мышца или группа мышц медленно вытягивается до максимальной, но комфортной точки. Эта точка держивается без пружинящих движений, чтобы запустить процесс обратного рефлекса вытяжения мышцы. В этом случае можно добиться большей амплитуды растяжки. Достигнув максимальной точки вытяжения мышцы, эта точка удерживается в течение 30 секунд. Таким образом можно добиться увеличения длины мышцы. Статичная растяжка может выполняться как активно, так и пассивно.
Это самый распространенный и рекомендуемый вид растяжки в лечебно-оздоровительном фитнесе.
ИЗОМЕТРИЧЕСКАЯ РАСТЯЖКА
Изометрическая растяжка – это вид статической растяжки с использованием сопротивления мышечных групп через изометрическое сокращение вытягиваемых мышц. Этот вид растяжки быстрее остальных помогает развить пассивную гибкость и является более эффективным, чем пассивная или активная растяжка по отдельности. Изометрическая растяжка также помогает развить силу мышцы под напряжением. Результаты опросов и исследований говорят о том, что изометрическая растяжка помогает уменьшить боль, которая может появиться после вытяжения мышц. Самые распространенные способы для создания необходимого сопротивления для выполнения изометрической растяжки – это создать желаемое сопротивление на вытягиваемую конечность, при помощи кого-то или чего-то (стены, пола, скамейки и пр.)
РАСТЯЖКА нвп
«Нервно-мышечное восстановление проприорецепции» – это техника, разработанная в конце 1940-х и начале 1950-х годов доктором медицинских наук Германом Кабатом совместно с физиотерапевтами Маргарет Нотт и Дороти Восс.
НВП сначала применялось для лечения больных параличом. Однако, Восс и Нотт вскоре поняли, что НВП можно использовать и в лечебной физкультуре. К 1954 году были проведены обучения и в 1956 году вышло первое издание Нервно-мышечного восстановления проприорецепции. С этого момента этот курс стал входить в программу обучения физиотерапевтов университетов.
В конце 1970-х годов НВП применялось для развития гибкости здоровых людей. С тех пор не только физиотерапевты, но и спортсмены считают НВП эффективным. Для достижения определенных результатов, которые необходимо было достичь определенным группам клиентов, были разработаны вариации НВП. Сейчас здоровые люди понимают, что при помощи НВП можно добиться быстрых результатов в растяжке.
Нервно-мышечное восстановление проприорецепции – это терапевтическая техника, которая выполняется с помощью партнера. Такой вид растяжки выполняется на изометрической фазе мышечного сокращения. Растяжка удерживается 8 секунд, затем положение ослабляется и снова выполняется растяжка, но уже с большим усилием. Целью этой растяжки является препятствие рефлексу растяжения мышц, то есть развитие гибкости.
ДЫХАНИЕ
В программе «СПИРАЛИ» очень важно соотносить дыхание и движение. В «СПИРАЛЯХ» используется грудное боковое дыхание, традиционное для Пилатес. Ниже с целью подробного описания механизма дыхания приведено описание дыхательной системы.
ДЫХАТЕЛЬНАЯ СИСТЕМА
ЛЕГКИЕ
Легкие выглядят как два эластичных мешочка, лежащих в грудной клетке, недалеко от сердца. Воздух поступает в легкие через жесткую трахею (дыхательное горло), которая открывается неполными кольцами хряща. Трахея затем разделяется на два бронха, каждый из которых находится в легких. Затем бронхи делятся на более мелкие ветви, которые называются бронхиолами, и затем на еще меньшие ветви, которые заканчиваются маленькими воздушными мешочками – альвеолами. Эти маленькие воздушные мешочки делают структуру легких губчатой, что позволяет пространству, внутри которого находятся газы, увеличиваться в тысячи раз.
Каждая альвеола настолько мала, что толщина ее стенки равна одной клетке. Сеть капилляров служит каждой альвеоле, так как газообмен происходит через тонкие стенки альвеол и капилляров.
ГАЗООБМЕН
Организм не любит дисбаланс. Например, в главе «Механика дыхания» описывается, как давление воздуха внутри и снаружи выравнивается в процессе дыхания. Содержание газа внутри альвеол и в крови отличается, и в процессе диффузии в организме восстанавливается баланс.
Вдыхаемый в альвеолы воздух имеет больший процент СПИРАЛЕЙ, чем кровь в капиллярах. Тонкие стенки альвеол и капилляров пропускают СПИРАЛИ, которые затем проникает в кровь. Здесь в красных кровяных тельцах СПИРАЛИ соединяются с гемоглобином и начинает свой путь обратно к сердцу. И наоборот, содержание углекислого газа в крови в капиллярах выше, чем в воздухе в альвеолах, и поэтому углекислый газ проходит через стенки клеток и в легкие, где воздух выдыхается. Этот процесс называется газообменом.
МЕХАНИКА ДЫХАНИЯ
Объем полости грудной клетки увеличивается и уменьшается, когда воздух попадает в легкие и выходит из них.
Увеличение объема полости грудной клетки приводит к снижению давления внутри нее. В этот момент воздух снаружи тела находится под большим давлением, чем в полости грудной клетки. Это более высокое давление направляет воздух в легкие, заставляя их расширяться. Мышцы, отвечающие за увеличение размера полости грудной клетки, затем расслабляются, позволяя ей тем самым уменьшаться в размере и выводить воздух из легких.
Процесс вдыхания называется инспирацией, а выдыхания – экспирацией.
infopedia.su